
research article | DOI: https://doi.org/10.31579/2690-8816/075
1 R. S. Mehta Jain Department of Biochemistry and Cell Biology, Vision Research Foundation, Sankara Nethralaya, Chennai – 600 006, India.
2 Department of Uveitis, Medical Research Foundation, Sankara Nethralaya, Chennai – 600 006, India.
*Corresponding Author: M. Bharathselvi, R.S. Mehta Jain Department of Biochemistry and cell Biology, Vision Research Foundation, Sankara Nethralaya, 41, College Road, Chennai-600 006.
Citation: Muthuve B, Konerirajapuram N. S, Ramanujam S, Jyothirmay B. (2022). Inflammation mediated alterations in the iron homeostasis of Eales’ Disease. J. Clinical Research Notes. 3(5); DOI: 10.31579/2690-8816/075
Copyright: © 2022 M. Bharathselvi, this is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: 27 June 2022 | Accepted: 04 July 2022 | Published: 20 July 2022
Keywords: eales’ disease; idiopathic retinal vascular disease; iron transport; hepcidin
Background: Eales’ Disease is an idiopathic retinal periphlebitis, characterized by recurrent vitreous hemorrhage, neovascularization and inflammation. The disease distresses the retina of adult males between 15 and 45 years. In the present study proteins involved in iron homeostasis were assessed in serum and peripheral blood mononuclear cells.
Methods: Forty male subjects, were recruited for the study. Their blood samples were used to measure the ferritin, transferrin, soluble transferrin receptor, hepcidin, ferroportin and heme. Besides ALAS, HO, HIF-2 and other relevant parameters were also measured.
Results: In the ED group, significantly increased in the levels of heme, heme oxygenase, ferritin and VEGF were observed in serum and monocytes of ED, besides decrease in the levels of transferrin. Interestingly, the expression levels of hepcidin and HIF were increased whereas the ferroportin was found to be decreased.
Conclusions: These results propose evidence for the involvement of altered iron homeostasis.
Eales’ disease (ED), characterized as an idiopathic, inflammatory veno-occlusive disease, which affects, the peripheral retinal veins especially of young males [1,2]. The disease is found to be, prevalent in Indian subcontinent with an incidence rate of 1 in 200-250 ophthalmic patients [3,5]. ED presents with periphlebitis that causes retinal ischemia and persistent ischemia leads to retinal neovascularization and consequential blindness [6]. Although, ED is associated with multiple causes, its etiopathogenesis is not clearly known, till date [7]. Among various causes, notable changes in iron homeostasis were observed in patients with ED viz., increased ferric (Fe3+) to ferrous (Fe2+) ion ratio in the serum and increased levels of iron in vitreous and monocytes, of patients with ED [8,9] . Apart from ED; iron is also, involved in various other ocular diseases, like age related macular degeneration (AMD), cataract, glaucoma conditions which results in intraocular hemorrhage [10]. Moreover, iron is a potent producer of most reactive hydroxyl radicals and can be the reason, for the considerable oxidative stress. Oxidative stress, hydroxyl radicals and decreased antioxidants levels were reported in vitreous, serum and monocytes obtained from ED patients [9-11]. The increased level of advanced glycation end product - carboxy methyl lysine also, confirms the involvement of iron induced glycoxidation in Eales' disease [12]. Further a novel 88 kDa proteins, present in serum and vitreous of ED patients, characterized to be a glycoprotein with iron binding capacity and antioxidant role [12].
Dietary iron; imported by the DMT1 (Divalent metal transporter-1) which is expressed on the apical side of absorptive cells, is reduced from Fe3+ to Fe2+ form by duodenal cytochrome B [10]. The iron thus, imported is either stored in cytosolic iron-storage molecule ferritin or secreted into plasma by the basolateral iron exporter ferroportin [13]. The plasma enzymes ceruloplasmin and hephaestin reconvert Fe2+ to Fe3+ that bind to transferrin and transferred to heme group. In case of iron overload; hepcidin, a regulatory protein of iron absorption, triggers the degradation of ferroportin, thereby, preventing release of excess iron into the circulation [14,15].
It is noteworthy that, hepcidin, a key regulator of iron homeostasis [15]. is induced as a result of inflammation, a key factor involved in the pathogenesis of ED. To understand the relevance of iron homeostasis and its deviation in pathophysiology of ED, there exists a need to decipher the functionality of proteins involved in iron metabolism.Therefore, the study aims to unravel the derangement in iron metabolism in ED patients, by using serum and PBMCs from case and control subjects and this study, is first of its kind to provide insights in the understanding of the iron-mediated etiopathology of the disease.
Patients
Twenty patients with ED, of unknown etiology (as diagnosed after detailed fundus examination by an ophthalmologist), and twenty healthy adult volunteers were recruited for the study. ED was diagnosed based on clinical features such as vasculitis, vitreous hemorrhage, peribhlebitis, vascular sheathing, peripheral nonperfusion, presence of floaters and decrease in visual acuity. The demographic details of the patients are given in table 1. All forty test subjects were males, between the ages of 15 and 45, and were generally healthy. None of them were smokers or alcoholics or taking vitamin supplements. The authors’ Institute’s Research Board and Ethics Committee approved this study. Informed written consent for participating in the study was obtained from all participants. All experiments pertaining to human subjects were performed in adherence to the tenets of the Helsinki declaration.
S. NO | Age/Sex | Clinical Diagnosis | Medications at time of collection |
Patients | |||
1 2 3 4 5 6 7
8 9 10 11
12 13
14 15 16 17 18 19 20 | 25/M 20/M 21/M 26/M 20/M 43/M 18/M
32/M 42/M 33/M 23/M
30/M 42/M
28/M 15/M 23/M 43/M 17/M 30/M 17/M | active vasculitis, cotton wool spot (OD,OS) active vasculitis, vitreous hemorrhage active vasculitis, vitreous hemorrhage OD-perivasculitis,choroiditis active vasculitis,vitreous hemorrhage,ED Vasculitis Active vasculitis, dispersed vitreous hemorrhage active vasculitis Vitreous hemorrhage, healed vasculitis Vasculitis, mild vitreous hemorrhage active vasculitis, neovascularization active vasculitis, vitreous hemorrhage, Bronchial tuberculosis active vasculitis, vitreous hemorrhage peripheralvasculitis,vitreous hemorrhage,phlebitis, healed vasculitis active vasculitis, subhyaloid hemorrhage active vasculitis,vitreous exudates, ED active vasculitis,vitreous hemorrhage, active vasculitis,hemorrhage active vasculitis,vitreous hemorrhage activevasculitis,vitreous hemorrhage, periphlebitis | Nil Nil Nil Nil Nil Nil Nil
Nil Nil Nil Nil
Nil Nil
Nil Nil Nil Nil, prednisone Nil Nil Nil |
Controls | |||
|
| 20 healthy volunteers with the age group of 27.5 ± 0.9 | Nil |
Table 1: Demographic details of the patients
Sample collection
From each participant, 12 ml of blood sample was collected. In the 4 ml blood sample was collected in plain tubes and serum was separated from blood cells by centrifugation at 3000 g at 25ºC for 10 minutes, aliquoted and stored at -80ºC for further analysis. 8 ml of heparinized blood sample was collected and monocytes were isolated by ficoll density gradient centrifugation.
Biochemical parameters analyzed in serum sample were as follows; heme, aminolevulinic acid synthase, heme oxygenase, ferritin, transferrin, soluble transferrin receptor and VEGF by following various methods described below. Parameters analysed in monocytes included ferritin, heme, heme oxygenase, aminolevulinic acid synthase and VEGF. RNA was isolated and mRNA expression of hepcidin, ferroportin, and HIF2α were quantified by Q-PCR.
Determination of Heme, Heme oxygenase activity (HO) and Aminolevulinic acid synthase (ALAS)
Heme content in serum and monocytes was determined using colorimetric determination at 400 nm by using a QuantiChrom Heme assay kit described by Berry et al. [16] (Bio Assay Systems, Hayward, CA, USA). The concentration of heme was expressed as µM in serum and µm / mg of protein in monocytes. HO activity assay in serum and monocytes was performed by using the method described by Bussolati et al., [17]. The specific activity of HO was expressed as pmoles of bilirubin formed / mg of protein / h in serum and monocytes [17]. Aminolevulinic acid synthase activity in serum and monocytes was performed by the method described by Hunter et al [18]. The concentration of ALAS was expressed as U/L in serum and units/mg protein in monocytes.
Determination of Ferritin, Transferrin and soluble transferrin receptor
Ferritin in serum and monocytes were determined by turbidimetry method described by Bernard [19]. The concentration of ferritin in serum and monocytes were expressed as µg/L in serum and in monocytes µg/mg of protein. Transferrin in serum was determined by turbidimetry method described by Kreutzer [20]. and the concentration of transferrin in serum was expressed as mg/dL and soluble transferrin receptor was performed by the method of enzyme linked immunosorbent assay described by Samuelson [21] and the concentration of serum transferrin receptor in serum was expressed as mg/L (ELISA, BioVendor, Canada).
Determination of Vascular endothelial Growth Factor (VEGF)
VEGF in serum and monocytes was determined by using Quantikine VEGF Immunoassay kit (R&D, USA) in accordance with the manufacturer’s instructions. The concentration of VEGF in serum was expressed as pg/mL and in monocytes was expressed as pg/mg of proteins. The optical density of well was determined using a microplate reader at 450 nm.
Isolation of total RNA, reverse transcriptase and real-time PCR assays
RNA was extracted by using TRIZOL reagent (sigma), cDNA quality and concentration were estimated by optical density using Nanodrop1000 (Thermo Scientific, Wilmington, DE). Quantitative real-time PCR assays were developed using RNA Master SYBR Green Kit according to the manufacture’s protocol using Applied Bio systems [22] (Foster City, CA). Levels of hepcidin, ferroportin and hypoxia inducible factor - HIF 2α were determined and the details of primers given in table 2.
S.NO | Gene Name | Accession number | Forward primer | Reverse primer |
1 | Hepcidin | 5′GACACCCACTTCCCCATCTG3′ | 5′ GCAGGGCAGGTAGGTTCTAC3′ | |
2 | Ferroportin | NM_014585.5 | 5′ TTGGGGAGATCGGATGTGGC 3′ | 5′ GTCACCGATGATGGCTCCCA 3′ |
3 | HIF 2 α | NM_001430.4 | 5′AGGTGGAGCTAACAGGACATAG3′ | 5′GCTGACTTGAGGTTGACAGTAC3′ |
4 | GAPDH | NM_002046 | 5′GCCAAGGTCATCCATGACAAC 3′ | 5′GTCCACCACCCTGTTGCTGTA 3′ |
Table 2: RT- PCR primers
Statistics
All values were expressed as mean ± standard error mean. With SPSS software, (version: 16.0) the raw data were analyzed for statistical significance using student “t” test. P value of < 0>
The 2 –ΔΔCt method was used to estimate relative transcript levels with GAPDH as endogenous control. Relative quantification data were obtained using ABI prism 7000 SDS software. The PCR reactions were set in triplicate, the mean values used for calculations.
Results
Increased levels of ALAS, Heme, Heme oxygenase and VEGF
The serum and intracellular levels of ALAS, Heme, Heme oxygenase and VEGF are given in table 3 and figure 1. Since serum haemoglobin was more in ED, may be of excess production or breakdown of haemoglobin. To address this, we measured ALAS. There was a significant increased levels of serum ALAS, Heme, HO and VEGF in ED patients compared to the control subjects (p < 0 xss=removed xss=removed xss=removed xss=removed xss=removed xss=removed>
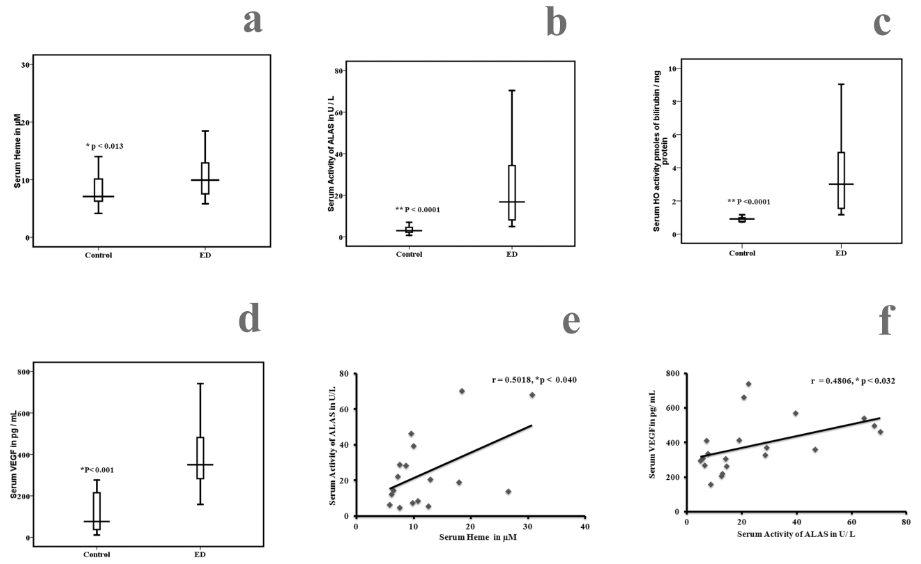
Figure 1: Comparison of serum Heme, aminolevulinic acid synthase, Heme oxygenase (HO) and VEGF among ED and healthy controls. Statistical significance between control (n = 20) and ED
(n = 20), a) Heme p < 0>
Correlation between serum aminolevulinic acid synthase, serum Heme, and serum VEGF in ED and controls, e) Positive correlation between serum Heme and aminolevulinic acid synthase (r = 0.50018, p < 0 xss=removed>
Levels of Ferritin, Transferrin and Soluble transferrin receptor
We measured the levels of serum ferritin, serum transferrin and soluble transferrin receptor levels in study subjects. The results are given in figure 2. There was a significant increase in the levels of serum and intracellular ferritin (table 3) in ED patients compared to the controls (p < 0 xss=removed>
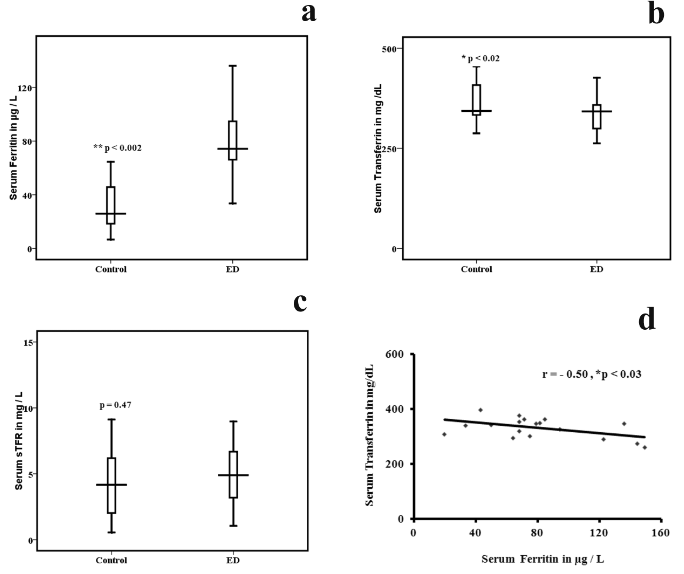
Figure 2: Comparison of serum ferritin, Transferrin and soluble transferrin receptor among ED and healthy controls. Statistical significance between control (n = 20) and ED (n = 20),
a) Serum ferritin p < 0 xss=removed>
d) Negative correlation between serum ferritin and serum Transferrin in ED and controls,
(r = -0.50, p < 0>
Expression of Iron regulators
Why there is an increased intracellular ferritin levels, in ED subjects, provoking interest in knowing the status of iron regulators, their role in the disease and hence to measure the mRNA expression of hepcidin an iron sensor and ferroportin an iron exporter. The mRNA expression of hepcidin was up regulated and ferroportin was down regulated in ED patients when compared to the controls (figure 3). Thus, the result shows a potential role for that hepcidin in ED. This increase in the expression of hepcidin may be due to increased iron, inflammation or infection. It is known that hepcidin can bind to ferroportin, triggering its internalization and degradation resulting in the accumulation of intracellular iron in the form of ferritin.
Over expression of Hypoxia inducible factor (HIF2 α)
Since, both VEGF and HO are elevated in ED; it is of interest to know further on the status of hypoxia inducible proteins. Hence, the expressions of hypoxia (HIF 2) were measured. The mRNA expression of HIF 2 was up regulated in ED compared to the controls as shown (figure 3).
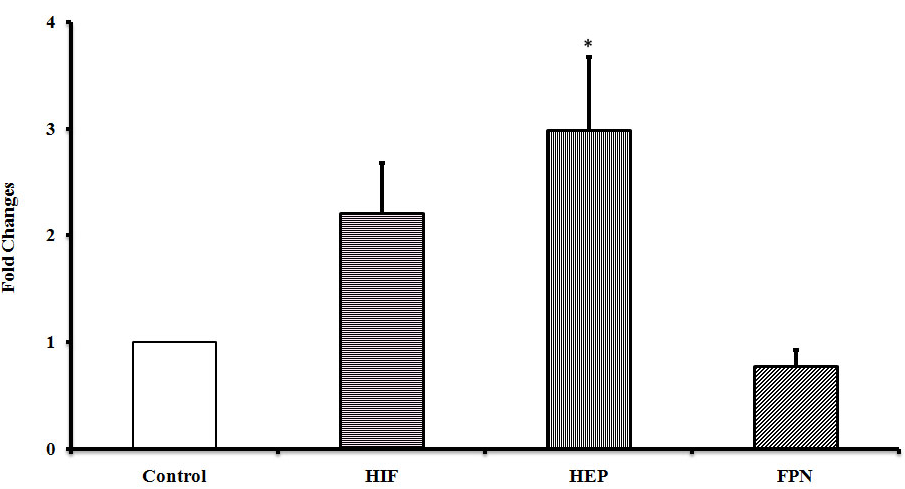
Figure 3: mRNA expressions in human PBMC among ED and healthy controls. Error bars represents SE and significance represents * of the relative expression levels normalized against GAPDH.
The aetiopathogenesis of ED is not clearly known. However, recently literature associated ED with tuberculosis, but still now the active organism was not cultivated. The disease still remains idiopathic, with no diagnostic test available in the lab. Direct evidence of infection is not made. Similar to homocysteine, Iron also acts as an independent risk factor of cardiovascular diseases. Roest [23] observed increased iron stores leads to cardiovascular diseases similar to that of hyperhomocysteine. Sullivan [24] observed that increased iron stores lead to endothelial dysfunction and that elevated homocysteine is dependent on iron overload. This excess iron generates free hydroxyl radical and causes oxidative stress through Fenton reaction. Schiepers [25] observed that changes in both homocysteine and ferritin levels in cardiovascular disease. The serum ferritin is frequently used as a measure of iron stores. The present findings indicate iron storage since; there is an elevated level of ferritin and lowered transferrin in ED.
It is known that, red cell breakdown can occur outside or inside the vascular compartment in ED. Increased serum haemoglobin in ED indicates the vascular damage due to intra vascular and extra vascular hemolysis. Extra vascular hemolysis is occurred due to phagocytosis whereas intravascular hemolysis as a result of mechanical injury and toxic factors. Heme is the prosthetic group of heme proteins such as haemoglobin, is an essential molecule plays a crucial role in cell differentiation and other functions. Free heme namely unbound heme can be toxic to cells, because it results in production of reactive oxygen species and causes cell damage. The absolute levels of Heme are regulated by its biosynthesis and catabolism. Heme biosynthesis is regulated by ALAS and its degradation to Fe, bilirubin and CO is catalyzed by HO, in this study, it is observed that both the enzymes are elevated in ED.
Heme oxygenase plays an important role in regulating the heme level by catalyzing the initial and rate limiting step of heme degradation and resulting in the formation of carbon monoxide, iron and bilirubin. Heme oxygenase exists as three isoforms; HO-1, HO-2 and HO-3. HO-1, the inducible 32-kDa isoform, HO-2, the constitutive 36-kDa isoform, and HO-3, has no activity and is not expressed in humans. The HO-1 is a member of the heat shock proteins, and its expression is influenced by hypoxia [26] heavy metals, ROS such as H2O2 [27], reactive nitrogen oxides [28], TNF α, interleukin β and interferon ϒ [29]. The biological functions of HO-1 are associated against oxidative and cellular stress. HO-1 represents a crucial mediator of antioxidants and possesses anti-inflammatory and anti- apoptotic properties [30]. L’Abbate et al., and Bharathselvi et al.,[31, 32] have shown that induction of HO-1 was associated with a parallel increase in the serum levels of adiponectin, which has a well-documented anti-inflammatory property. The peroxisome proliferator-activated receptor (PPAR-ϒ) regulates the expression of HO in human vascular cells [33].
Over expression of HO-1 contributes in the revascularization of damaged tissue. In term of neovascularization, HO-1 having a pro-angiogenic, anti-inflammatory and anti-apoptotic enzyme in regulation of wound healing [34,35]. Product of HO activity the bilirubin is a powerful antioxidant thereby, protecting the retinal cells. However, during haemorrhage, the iron and bilirubin excessively produced and are neurotoxic, have deleterious consequences. HO-1 is an inducible enzyme whose activity increases in response to iron as well as heme, light, oxidative stress, and inflammation. The main condition for the initiation of neovascularization is hypoxia [36]. Hypoxia-inducible factor activates several genes related to iron metabolism such as HO-1, endothelin-1, transferrin, transferrin receptor and ceruloplasmin [36].
HO, cleavage of the heme ring will release intracellular iron, which in turn increases the stimulation of ferritin. Dulak et.al observed that HO plays an important role in angiogenesis during hypoxia, similarly nitric oxide synthase, by VEGF production [37]. Alternately VEGF can stimulate HO-1 to promote angiogenesis and inhibiting leukocyte adhesion and transmigration [17]. In the present study, it’s observed that both HO and VEGF increased due to inflammation and non-inflammatory responses.
Hepcidin, a circulating peptide hormone is mainly synthesized by the liver hepatocytes, and also in eye plays a major role in regulating iron homeostasis in the body [38]. The mature form is 25 amino acids with four inter subunit disulfide bonds. The massive iron overload found in hepcidin knock-out mice suggests that hepcidin is an iron stores regulator involved in communication of body iron status to the intestine and also in the retinal pigment epithelial cells [38]. The mechanism of hepcidin activity depends on hepcidin interactions with ferroportin. Ferroportin is the only known mammalian cellular iron exporter. Hepcidin regulates post translational ferroportin expression [38]. Hepcidin binds to ferroportin and causes its internalization and degradation in turn blocks the iron transport via ferroportin [38]. Hepcidin is reported to be elevated in chronic inflammation condition, anemia, more specifically wherein iron accumulates inside the cells [39].
Hepcidin synthesized can be induced by inflammatory cytokine IL-6, inflammation, and infection (40)[Prentice, 2012 #102]. IL‐6 acts via its receptor and causes phosphorylation of signal transducer and activator of transcription 3 (STAT 3), STAT3 activation requires the presence of SMAD 4 to affect the HAMP gene expression [39]. Under normal conditions HAMP gene expression is regulated by BMP/SMAD and STAT3 pathways. Another hepatocyte iron sensors activating hepcidin synthesis are hemochromatosis protein (HFE) and transferrin receptor 2 (TfR2) [39].
Due to an intracellular storage of iron, there is an increased production of hepcidin. In the present study, there is an increased storage of ferritin levels and up regulation of hepcidin. Inflammation leads to cellular iron sequestering through IL-6 up regulation of hepcidin. Iron can be export from the cell with the help of iron exporter ferroportin. In the present study, we also observed that hepcidin expression was increased and ferroportin was decreased in ED compared to controls. Interestingly cellular iron accumulation caused diminished ferroportin after hepcidin binding. There is an increased expression of hepcidin may be due to increased iron, inflammation and infection. Importantly, a cellular iron act as a cofactor of HIF, in the present study, it shows that there is an increased expression of HIF2α. A previous report says that hypoxia is a negative regulator of hepcidin expression [39], but in the present study, shows that HIF 2α expression was increased in ED may be due to inflammation and also by IL-6. The interaction between hepcidin and ferroportin binding is a key step to control an iron homeostasis.
From the present study, we conclude that increased serum and intracellular ferritin, heme and HO, hepcidin play an important role in the vasculitis. The present study establishes the role for iron in disease pathogen. In our study thus the increased levels of these proteins may be due to an infection and inflammation conditions. The real significance of these findings needs to be understood in an animal model or an in-vitro cell culture experiment.
Biochemical Parameters | Control subjects (n = 20) | Eales’ disease (n = 20) | P value |
Ferritin (µg / mg of proteins) | 20.9 ± 1.4 | 65.7 ± 2.5 | 0.001 |
Heme (µm / mg of proteins) | 8.9 ± 1.7 | 27.2 ± 5.1 | 0.001 |
Heme oxygenase (pmoles of bilirubin / mg protein) | 1.5 ± 0.2 | 6.4 ± 0.5 | 0.001 |
Aminolevulinic acid synthase (U / mg of proteins) | 2.4 ± 0.2 | 12.5 ± 3.0 | 0.001 |
Vascular endothelial growth factor (pg/mg of proteins) | 29.8 ± 4.0 | 147.5 ± 40.2 | 0.005 |
Table 3: Intracellular Levels of Biochemical parameters measured in control and Eales’ disease subjects.
ED - Eales’ Disease
Hcy - Homocysteine
Hb - Haemoglobin;
HO - Heme oxygenase;
VEGF- Vascular endothelial growth factor;
ALAS - aminolevulinic acid Synthase;
IL - Interleukin;
Hep - Hepcidin;
FPN - Ferroportin.
Ethical approval: Institutional Ethics committee.
Consent to participate: Informed written consent for participating in the study was obtained from all participants. All experiments pertaining to human subjects were performed in adherence to the tenets of the Helsinki declaration.
Availability of data and materials: Data and materials are in the manuscript.
Competing Interests: No conflict of interest.
Funding: Supported by Grant from (ICMR) Indian council of medical research – ICMR/53/18/2009-BMS.
Authors’ Contributions: MB processing the sample analyzed the data and wrote the paper, RS processing the sample; KNS conceived the work, analyzed the data and wrote the paper. JB was the ophthalmic consultant who treated the patients, obtained their consent, recruited and in filling the clinical proforma in this study.
Acknowledgement: We thank the Indian Council of Medical research (ICMR) for their funding.
Authors’ Information (Optional): Not applicable
It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,

Dear Maria Emerson, Editorial Coordinator of International Journal of Clinical Case Reports and Reviews, What distinguishes International Journal of Clinical Case Report and Review is not only the scientific rigor of its publications, but the intellectual climate in which research is evaluated. The submission process is refreshingly free of unnecessary formal barriers and bureaucratic rituals that often complicate academic publishing without adding real value. The peer-review system is demanding yet constructive, guided by genuine scientific dialogue rather than hierarchical or authoritarian attitudes. Reviewers act as collaborators in improving the manuscript, not as gatekeepers imposing arbitrary standards. This journal offers a rare balance: high methodological standards combined with a respectful, transparent, and supportive editorial approach. In an era where publishing can feel more burdensome than research itself, this platform restores the original purpose of peer review — to refine ideas, not to obstruct them Prof. Perlat Kapisyzi, FCCP PULMONOLOGIST AND THORACIC IMAGING.
