
Research Article | DOI: https://doi.org/10.31579/2692-9406/008
*Corresponding Author: Samad Farashi Bonab, Department of Immunology, School of Medicine, Tehran University of Medical Sciences, Tehran, Iran.
Citation: Samad Farashi B, Maryam M. (2020) Differential expression of apoptosis related genes in the peripheral blood mononuclear cells of acute lymphoblastic/lymphocytic leukemia (ALL) patients. Biomedical Research and Clinical Reviews. 1(4); DOI: 10.31579/2692-9406/008
Copyright: © 2020 Samad Farashi Bonab, This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: 11 June 2020 | Accepted: 04 November 2020 | Published: 09 November 2020
Keywords: acute lymphoblastic/lymphocytic leukemia; pbmc, apoptosis; gene expression
Apoptosis is the cell’s intrinsic death program which plays a crucial role in the regulation of many normal physiological processes in the body’s tissues. In leukemia patients, the extent of cancer cell susceptibility to apoptosis is correlated with clinical responses to chemotherapy and disease prognosis. The aim of this study was evaluation of the expression of apoptosis related genes in the peripheral blood mononuclear cells (PBMCs) of acute lymphoblastic/lymphocytic leukemia (ALL) patients. Peripheral blood samples were obtained from 20 patients with ALL and 20 healthy individuals. PBMCs were isolated using Ficoll-Paque density gradient centrifugation method. After RNA extraction and cDNA synthesis, gene expression levels of apoptosis related genes including caspase-3, 8, 9, BAX, and BAK genes were measured by real-time RT-PCR technique. Gene expression analysis showed that the expression levels of the initiator caspases-8 and -9 are increased in the PBMCs of adult ALL patients when compared with that in PBMCs of healthy individuals. Increased gene expression levels of the proapoptotic protein BAK was also detected in PBMCs of ALL patients. In contrast, decreased expression levels of the proapoptotic BAX and the executioner caspase-3 were observed in the PBMCs of ALL patients. These results suggest that the expression of genes involved in both extrinsic and intrinsic signaling pathways of apoptosis are induced in PBMCs of ALL patients while the gene expression of other proapoptotic molecule, BAK, and the executioner caspase-3 diminished in PBMCs cells of ALL patients. This findings indicate that resistance to apoptosis may be one of the hallmarks of ALL cells.
Acute lymphoblastic/lymphocytic leukemia (ALL) is a malignancy of B or T lymphoblasts characterized by uncontrolled proliferation of abnormal, immature lymphocytes in the bone marrow, blood and extramedullary sites. Approximately 85% of childhood ALL cases and 75% of adult ALL cases develop from B cell precursors, while the remaining ALL cases develop from T cell precursors [1]. Disease symptoms arise from various degrees of anemia, neutropenia, and thrombocytopenia or infiltration of ALL cells into tissues. Leukemia cells in the peripheral blood can enter various organs such as the lymph nodes, spleen, liver, central nervous system, and skin [2]. ALL is the most common type of childhood leukemia, accounts for 75% of childhood leukemias [3] and makes up 25% of all childhood cancers. About 54% of ALL cases are diagnosed in patients aged younger than 20 years [4]. In adults, ALL is accounts for 20% of adult leukemia [5], and makes up 0.4% of all adult cancers. Adult ALL often occurs around the age 50, especially in those with history of chemotherapy, exposure to ionizing radiation, pesticides, certain chemicals or viruses [1]. In 2020, the estimated numbers of new cases of ALL and its related deaths in the United States are 6,150 and 1,520, respectively [6].Intensive chemotherapy can induce complete responses in ALL patients. However, survival rate for older patients achieving a remission is relatively low [7]. The 5‐year relative survival rate is 89% for ages less than 20 years, 47% for ages 20-49 years, 28% for ages 50-64 years, and 17% for ages 65 years and older [8].
It has been shown that the extent of cancer cell susceptibility to apoptosis is correlated with clinical responses to chemotherapy and disease prognosis [9, 10]. Apoptosis is a major form of cell death, characterized by a series of distinct morphological changes including DNA fragmentation, nuclear condensation and fragmentation, fragmentation of cytoplasmic organelles (the Golgi apparatus, endoplasmic reticulum and mitochondrial network), cytoskeletal rearrangement, plasma membrane blebbing, and cell shrinkage [11, 12]. These events are mainly orchestrated by members of a family of cysteine proteases known as caspases [13]. Caspase-dependent proteolysis leads to detachment of the apoptotic cell from the extracellular matrix and fragmentation of the genomic DNA, Golgi, endoplasmic reticulum and mitochondrial networks. Finally, the cell breaks into small membrane-surrounded fragments, known as apoptotic bodies, which are rapidly removed by phagocytes, a process that minimizes damage and disruption to neighboring cells and avoids activation of immune responses.
Caspases exist in an inactive form in the absence of an activation signal. Three major apoptotic pathways by which caspases can be activated have been identified, including extrinsic (or death receptor) pathway, intrinsic (or mitochondrial-mediated) pathway, and granzyme B pathway. Caspase-8 and -9 are the initiator (or apical) caspases in the extrinsic and intrinsic pathways, respectively. These caspases possess a protein interaction prodomain, which plays a role in their recruitment and subsequent activation within cellular complexes known as the death-inducing signaling complex (DISC) and the Apaf-1 apoptosome in the extrinsic and intrinsic pathways, respectively. Activated caspase-8 and -9 then process and activate effector caspases (also called executioner caspases) including caspase-3, -6, and-7. Granzyme B, a protease released from cytotoxic T cells and natural killer cells, initiates apoptosis by cleaving the proapoptotic BID protein as well as caspase-3 and -7 in the target cells [12]. Caspase-3 is the major effector caspase in both the extrinsic and intrinsic pathway as well as in the granzyme B pathway and is responsible for cleavage of multiple cellular substrates during apoptotic cell death.
The intrinsic pathway of apoptosis can be induced in cancer cells by various stimuli, including increase in the levels of pore-forming proapoptotic BCL-2 (B cell lymphoma-2) family proteins leading to pro-caspase-9 activation [14]. Mitochondrial outer membrane permeabilization (MOMP) is involved in this pathway of apoptosis. MOMP is controlled by members of the BCL-2 protein family. Members of this protein family are divided into three classes based on their sequence homology and function. The first class includes the antiapoptotic proteins BCL-2, BCL-XL, BCL-w, BCL-B, BCLA1, and MCL-1 which contain all BCL-2 homology domains (BH1-4). The second class includes the proapoptotic BH3-only proteins BID, BIM, BAD, BIK, NOXA, PUMA, HRK, and BMF. The third class contains proapoptotic effector proteins BAX, BAK, and BOK/MTD. If the proapoptotic members of the BCL-2 family overwhelm the antiapoptotic members, the mitochondrial-mediated pathway of apoptosis triggers and the cell dies. BAX (BCL-2-associated X protein) and BAK (BCL-2 antagonist/killer 1) are core regulators of the mitochondrial pathway of apoptosis. Once activated, BAX and BAK proteins homo-oligomerize and directly cause MOMP (by making pores in mitochondrial outer membranes) which results in efflux of mitochondrial intermembrane space proteins, such as cytochrome c, into the cytosol. Cytochrome c in turn seeds the multimolecular apoptosome complex assembly and activation of caspase-9.
The ability of cancer cells to evade apoptosis is one of the hallmarks of cancer [15]. Expression of apoptosis-related genes may affect cancer outcome and prognosis. The aim of this study was evaluation of the expression profile of some apoptosis-related genes in the peripheral blood mononuclear cells of ALL patients.
Sample collection from patients with leukemia
Peripheral blood samples were obtained during routine clinical assessment of 20 patients with ALL between 2017 and 2018. All the patients were referred to the Department of Hematology and Oncology, Shariati's Hospital of Tehran University of Medical Sciences, Tehran, Iran, for verifying the diagnosis. Diagnosis was based on standard clinical, morphological and immunophenotyping criteria. The general information of the patients was summarized in Table 1. Additionally, peripheral blood samples were obtained from 20 healthy donors without a history of any malignancies as controls. Informed consent was obtained from all subjects who participated in this study.
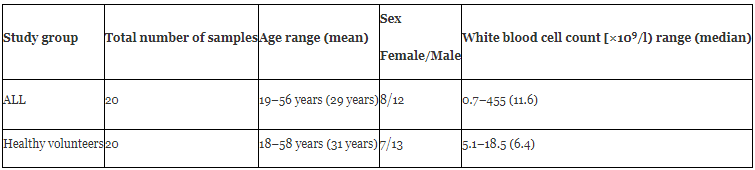
Isolation of peripheral blood mononuclear cells
Peripheral blood mononuclear cells (PBMCs) were isolated by density gradient centrifugation on Ficoll-Paque solution (Biosera, UK). Briefly, blood samples were diluted 1:1 with phosphate-buffered saline (PBS; Ca and Mg-free; Biosera, UK) and were subsequently layered onto the Ficoll-Paque in a centrifuge tube and centrifuged at 400 g for 30 min at 20°C. The PBMC layer was harvested and washed twice with PBS. Isolated PBMCs immediately subjected for the extraction of total RNA.
RNA extraction
For RNA extraction, PBMCs were lysed by adding 1 ml of Trizol reagent (Qiagen) and vortexing for 20 sec. The homogenized samples were incubated for 5 minutes at room temperature (RT) to permit the complete dissociation of nucleoprotein complexes. Then, 200 ul of chloroform was added to the samples, vortexed vigorously for 15 seconds, and incubated at RT for 2–3 minutes. Samples were then centrifuged at 12,000 g at 4°C for 15 min. Following centrifugation, the upper aqueous phase was carefully transferred into fresh RNase-free microtube. RNA was precipitated from the aqueous phase by mixing the aqueous phase with an equal volume of isopropanol and incubation at RT for 10 minutes followed by centrifugation at 14,000 g at 4°C for 10 min. The RNA pellet was washed by brief vortexing, mixing with 1 ml of 75% ethanol, and recentrifugated at 8,500 g at 4°C for 10 min. After removing leftover ethanol, the precipitated RNA was air-dried for 5-10 minutes and dissolved in 40 ul diethylpyrocarbonate (DEPC)-treated H2O. The quantity of extracted RNA was determined by measuring the absorbance at 260 nm and the RNA quality was assessed by determination of the ratio for absorbance at 260 nm vs. absorbance at 280 nm (A260 nm/A280 nm) using the Thermo Scientific™ NanoDrop™ One Spectrophotometer (Themo Fisher Scientific, USA). The RNA purity was also assessed by agarose gel electrophoresis. RNA samples had an A260/A280 ratio > 1.6. Contaminating DNA was removed from each RNA extract by means of DNase (Themo Fisher Scientific, USA) treatment, according to the manufacturer’s recommendations. RNA samples were stored at -80 °C.
cDNA synthesis
cDNA synthesis was performed on 1 µg of RNA in a 10 µl sample volume using High-Capacity cDNA reverse transcription kit (Applied Biosystems, USA) according to the manufacturer's instructions. Ten µl of a master mix containing 2 µl of 10X reverse transcriptase buffer, 0.8 µl of 25X dNTP mix (100 mM), 2 µl of 10 X reverse transcriptase random primers, 1 µl of MultiScribe™ reverse transcriptase, and 4.2 µl of nuclease-free H2O was added to the RNA sample. The final reaction mixture was incubated at 25°C for 10 min and 37°C for 120 min followed by 5 min at 85°C to inactivate the enzyme and then cooled to 4°C. cDNA was stored at −20°C.
Gene expression analysis by real-time reverse transcriptase polymerase chain reaction (RT-PCR)
Quantitative analysis of mRNA expression of apoptosis related genes was performed by real-time RT-PCR using a StepOnePlusTM Real-Time PCR System (Applied Biosystems, USA). Each reaction contained 7.5 µl of RealQ Plus 2x Master Mix Green High ROX™ (Ampliqon, Denmark), 1 µl of mixed forward and reverse primers (5 µM), 1 µl of cDNA, and 5.5 µl of DNase-free H2O. The sequences of primers were as follows: Forward, 5′-TGGTTCATCCAGTCGCTTTG-3′ and reverse, 5′-CATTCTGTTGCCACCTTTCG-3′ for capsae-3 gene; 5′-AGGAGCTGCTCTTCCGAATT-3′ and reverse, 5′-CCCTGCCTGGTGTCTGAAGT-3′ for caspase-8 gene; 5′-ACTAACAGGCAAGCAGCAAAGT-3 and reverse, 5′-ACATCACCAAATCCTCCAGAAC-3′ for caspase-9 gene; forward, 5′-GGTCCTGCTCAACTCTACCC-3′ and reverse, 5′-CCTGAGAGTCCAACTGCAAA-3′ for BAK gene; forward, 5′-GGGGACGAACTGGACAGTAA-3′ and reverse, 5′-CAGTTGAAGTTGCCGTCAGA-3′ for BAX gene; forward, 5′-AGGAATTCCCAGTAAGTGCG-3′ and reverse, 5′-GCCTCACTAAACCATCCAA-3′ for 18S rRNA gene. All reactions were prepared in duplicate. The PCR thermal program was 95°C for 15 min followed by 40 cycles of 95°C for 15 sec, 58°C for 30 sec, and 72°C for 30 sec. Then, a melting curve analysis was performed to confirm that a single PCR amplicon has been detected with the SYBR green. The relative expression levels of the target genes were normalized to that of the endogenous control gene, 18S rRNA. The data were analyzed using the comparative threshold cycle (2-ΔΔCT) method.
Statistical analysis
Data were expressed as mean ± SEM. Unpaired student’s t-test was used to analyze statistical differences between the means of gene expressions in ALL patients and healthy individuals (controls). P value of <0.05 was considered statistically significant. All analyses were performed using Graphpad Prism 8 software (GraphPad Software, Inc.).
Results
Two important characteristics (age, and sex distribution) of ALL patients were nearly similar to those of healthy subjects (Table 1). Of 20 patients, 16 (80%) patients had B-cell precursor ALL, and 4 (20%) patients had T-cell ALL.
Melting curve experiments were carried out after real-time RT-PCRs to exclude non-specific reactions (false-positives). The amplified products were treated with the following thermal program: heating at 95°C for 15 s, cooling to 60°C, holding at 60°C for 1 min, and then slowly heating again to 95°C in increments of 0.3°C. Melt curves were generated by continuous fluorescence acquisition in the temperature elevating period from 60 to 95°C. Melt curve analysis showed a single sharp park indicating formation of a single product.
For gene expression analysis, threshold cycle (CT) values were measured during the exponential phase of the PCR reaction. ΔCT was then calculated by the difference between the CT values of the Caspase-3, 8, and -9, BAX, BAK and the CT value of the reference gene, 18S rRNA. Finally, relative expressions were calculated by 2-ΔΔCT equation.
Analysis the mRNA levels of apoptosis related genes (Caspase-3, 8, 9, BAX, and BAK) in PBMCs showed that expression of some apoptosis related genes were significantly altered in PBMCs of ALL patients (Figure 1). The expression of the initiator caspase-8 and -9 genes were significantly upregulated while the expression of the proapoptotic BAX and the executioner caspase-3 genes were significantly downregulated in PBMCs of ALL patients as compared with PBMCs of healthy individuals (p< 0.0001 for each). The expression of the proapoptotic BAK gene was also upregulated in PBMCs of ALL patients as compared with PBMCs of healthy individuals (individuals (p< 0.01).
The mRNA levels of caspase-8, -9 and BAK in the PBMCs of leukemia patients were 2.43, 5.56, and 1.52 folds higher than that in PBMCs of healthy subjects, respectively. In contrast, the mRNA levels of BAX and caspase-3 in the PBMCs of leukemia patients were 2.3 and 1.72 folds lower than that in the PBMCs of healthy subjects.
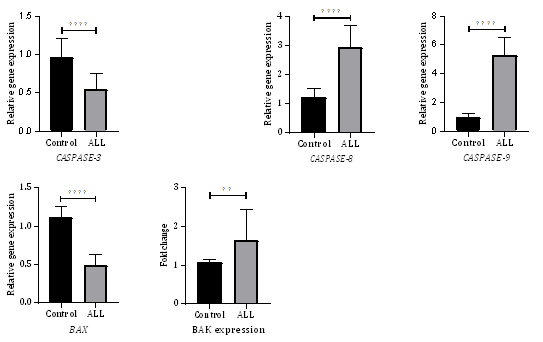
Caspase-8 is the initiator caspase in the extrinsic pathway of apoptosis. Caspase-8 is essential for induction of the extrinsic (the death receptor) apoptotic pathway. Activated caspase-8 initiates the proteolytic cleavage of caspase-3, -7 or BID. In certain cell types, known as Fas type I cells, the activation of caspase-8 is sufficient to induce the activation of downstream caspases such as caspase-3 [16]. In Fas type II cells, caspase-8 cleaves BID, leading to activation of the downstream components of the mitochondrial apoptotic pathway [17]. In some lymphoid cell lines, caspase-8 is vigorously activated and activates the downstream procaspases such as proscaspase-3 [18]. Caspase-9 is the initiator caspase in the intrinsic pathway of apoptosis and directly activates the effector caspase-3 and -7. Caspase-3 is at the crossroad of both the extrinsic and intrinsic pathways of apoptosis. Activated caspase-3 cleaves multiple cellular substrates involved in cellular integrity and metabolism which contribute to the cell death. Upregulation in caspase-3 gene expression ensures the presences of sufficient amounts of the proenzyme to eliminate apoptotic cells.
Low levels of caspases can be contributable to the decrease in apoptosis in cancer cells. Teitz and colleagues reported that the caspase-8 gene is frequently inactivated in childhood neuroblastomas [19]. Winter and colleagues found no significant differences in the mRNA levels of caspase-1, -3, and -9 in prostate tumors relative to the normal gland. But, protein expression analysis revealed that more than 80% of prostate tumors exhibit total lack of caspase-1 and reduced caspase-3 protein expression [20]. Devarajan and colleagues showed that about 75% of tumor and normal peritumoral tissue samples obtained from breast cancer patients lack caspase-3 mRNA and protein expression. Caspase-3 mRNA levels were also undetectable in commercially available total RNA samples isolated from breast and cervical tumors and were substantially decreased in RNA samples isolated from ovarian tumors. Protein expression levels of caspase-8 and -9 were normal in all of the caspase-3-negative breast tumor samples [21]. Fong and colleagues observed lower expression of caspase-8 and -10 mRNA and/or protein in choriocarcinoma and hydatidiform mole when compared with normal placentas [22]. Shen and colleagues found that the expression of caspase-9 mRNA is downregulated in stage II colorectal cancer tissues and correlates with poor clinical outcome [23]. The expression of caspases-2, 3, and 6 genes was heterogeneous in bone marrow samples obtained from patients with de novo acute myeloid leukemia (AML) (n=15). mRNA levels of caspase-3 were higher in most AML bone marrow samples compared to bone marrow samples obtained from healthy donors (n=2). Conversely, mRNA levels of the short isoform of caspase-2, that encodes an anti-apoptotic protein, were lower in most AML bone marrow samples than that in normal bone marrow cells [24]. In our study, mRNA levels of caspase-8, -9 were higher in the PBMCs of adult ALL patients compared to PBMCs of healthy individuals. In contrast, the mRNA levels of caspase-3 in the PBMCs of leukemia patients were lower than that in the PBMCs of healthy subjects. It is possible that cancer cells undergo apoptosis, recover, and proliferate if caspase-3 activation is inhibited.
BAX and BAK have crucial roles in the mitochondrial pathway of apoptosis. Dysregulation of BAX has been proposed to be involved in the survival of B cell chronic lymphocytic leukemia [25, 26]. Numerous other studies showed the role of BAX in leukemia progression as well as cancer cell resistance to apoptosis induced by chemotherapy or radiotherapy [27, 28]. Genomic loss of BAX has been observed in human colon cancers [29, 30]. Loss of BAX expression was also found in some other cancer types. Cartron and colleagues examined the expression of BAX and BAK in 50 human glioblastoma multiforme tumors and observed that all the tumors expressed BAK, while three did not express BAX. The levels of antiapoptotic proteins BCL-2 and BCL-XL or the BH3-only proapoptotic BID were approximately similar between BAX-expressing and BAX deficient tumor cells, while the levels of BAK were upregulated in BAX-deficient tumor cells [31]. Protein levels of BCL-2, MCL-1. BCL-XL, and BAX in pretreatment AML (n=123) and ALL (n=36) samples varied widely between different samples [32]. Low BCL-2 and high BAX protein levels were found in AML cells undergoing spontaneous apoptosis in vitro, whereas nonapoptotic AML cells showed high BCL2 and low BAX levels[33]. In another study, no clear correlation between both BCL-2 and BAX protein levels and the extent of spontaneous apoptosis were observed in adult AML cells in vitro [34]. Assessing the protein levels of BCL-2, BAX, MCL-1, and BCL-X in lymphoblasts from 47 childhood ALL showed that BCL-2, BAX, and MCL-1 were expressed in all samples. BCL-2 levels and BCL-2/BAX and MCL-1/BAX ratios were significantly higher in B lineage as compared with T-lineage disease. High BAX protein levels were significantly correlated with increased risk of relapse in childhood ALL [35]. Higher expression of BAX gene was found in 55 childhood primary B cell ALL patients when compared with their normal counterparts, B cell precursors [36]. BAX, BCL-2, and BCL-xL mRNA levels were significantly higher in 46 childhood ALL patients at their initial diagnosis than in 50 ALL patients in remission, while BAD mRNA levels were found to be lower in ALL patients than in remission patients [37]. In our study, mRNA levels of BAX in the PBMCs of adult ALL patients were lower than that in the normal PBMCs, while mRNA levels of BAK in the PBMCs of adult ALL patients were higher than that in PBMCs of healthy subjects. These results indicate downregulation of BAX gene expression and compensatory gene expression of the alternative mitochondrial pore-forming protein, BAK, in the PBMCs of ALL patients.
Although some differences in the levels of expression of apoptosis related genes in leukemia patients reported in various studies might be due to the fact that nonquantitative methods were used in some of that studies, overall results suggest that the expression of some apoptosis related genes is heterogeneous among leukemia patients. Nevertheless, it is now obvious that the gene expression and function of certain molecules that regulate or induce apoptosis are important factors in cancer cell survival and resistance to chemotherapeutic drugs. Induction of apoptosis is a complicated process and is tightly regulated by the activity of several genes. Furthermore, synthesis of new apoptosis related proteins is not required for induction of apoptosis. However, accumulation of zymogen forms of multiple caspases and BCL-2 family proteins through direct transcriptional mechanism occurs in cancer cells. Our results indicate that induction of apoptosis initiate in adult ALL cells, but cancer cells can block it. These findings suggest that choosing appropriate class of chemotherapy drugs based on alterations in apoptosis related genes such as BAX and caspase-3 genes may be beneficial to improve therapeutic efficacy of chemotherapy in leukemia patients.
Defects along apoptotic pathways can play a crucial role in carcinogenesis. Defects in apoptosis also play a key role in cancer cell resistance to some anticancer cytotoxic drugs. Regulation of apoptosis related genes transcription as well as compensatory expression of alternative apoptosis related genes affect the survival of cancer cells as well as the sensitivity of cancer cells to multiple apoptotic stimuli exploited by anticancer therapy. Reduced expression of apoptosis related genes such as the executioner caspase-3 and the proapoptotic effector BAX can be contributed to evasion of apoptosis. Further studies on the effects of these altered gene expressions in apoptosis-mediated cancer cell death may allow the identification of novel therapeutic targets or appropriate combinational therapy for ALL patients.
It was my pleasure to submit my testimonial concerning the Reviewer Board of our Scientific Journal “Brain and Neurological Disorders”. The Reviewers focused on some modifications and their contribution was helpful. The ladies of our Editorial Office were also supported my efforts. It was my honor to have such a co-operation and I am looking forward for more collaboration.

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Thank you for the speedy and efficient peer review process. I appreciate the fact that your peer reviewers do not take months to respond like with some other journals. I would also like to thank the editorial office for responding quickly to my questions. It is an excellent journal. I plan to submit more manuscripts in the future. Best wishes from, Robert W. McGee

Dear Grace Pierce, Editorial Coordinator of Journal of Clinical Research and Reports, Working with you and your team on our recent publication in JCRR has been a truly wonderful and enjoyable experience. The responses were prompt, and the reviewers were patient, constructive, and highly professional. One reviewer in particular gave me the feeling that a professor was carefully reading and commenting on my coursework, which was deeply touching. The entire process was straightforward and hassle‑free, with no tedious online forms to complete. I highly recommend this journal. Best wishes from, DR Aibing Rao, Head of R&D

I Appreciate the Opportunity to Share my Experience with the Journal of Clinical Research and Reports. The peer review process was timely and constructive, and the feedback provided helped improve the quality of our manuscript. The editorial office was professional, responsive, and supportive throughout the process, ensuring smooth communication and efficient handling of the submission. Overall, it was a positive experience collaborating with your team.

Dear Mercy Grace, Editorial Coordinator of Obstetrics Gynecology and Reproductive Sciences, We would like to express our gratitude for your help at all stages of publishing and editing the article. The editors of the magazine answer all the necessary questions and help at every stage. We will definitely continue to cooperate and publish other works in the Obstetrics Gynecology and Reproductive Sciences! Best wishes from, Alla Konstantinovna Politova,

Dear Maria Emerson, Editorial Coordinator of International Journal of Clinical Case Reports and Reviews, What distinguishes International Journal of Clinical Case Report and Review is not only the scientific rigor of its publications, but the intellectual climate in which research is evaluated. The submission process is refreshingly free of unnecessary formal barriers and bureaucratic rituals that often complicate academic publishing without adding real value. The peer-review system is demanding yet constructive, guided by genuine scientific dialogue rather than hierarchical or authoritarian attitudes. Reviewers act as collaborators in improving the manuscript, not as gatekeepers imposing arbitrary standards. This journal offers a rare balance: high methodological standards combined with a respectful, transparent, and supportive editorial approach. In an era where publishing can feel more burdensome than research itself, this platform restores the original purpose of peer review — to refine ideas, not to obstruct them Prof. Perlat Kapisyzi, FCCP PULMONOLOGIST AND THORACIC IMAGING.
